
为什么说CCR5变异基因只在北欧和西欧人中有

,欧洲人有是因为3000年前他们的一个老祖宗身上发生了基因变异,传下来的。就像是有特异功能的家族一样。
什么细胞表达CCR5,什么细胞表达CXCR4
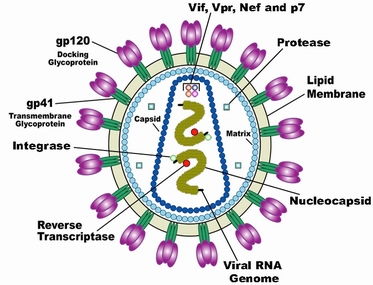
CCR5作为HIV?1受体拮抗剂的理想靶点,其拮抗剂药物抑制R5嗜性的HIV?1感染细胞的机制是,它们与CCR5结合后,使CCR5构象发生变化不利于gp120的识别或者导致CCR5的内源化作用(internalization),阻断了HIV?1与细胞包膜蛋白结合,导致HIV?1与CCR5在细胞表面结合的数量减少,从而起到抗感染作用。同时,有些学者担忧靶向于CCR5辅助受体的抗病毒治疗是否会加快R5嗜性病毒株向X4嗜性病毒株的转变。在体外实验中显示,除了一种耐药株产生嗜性转换外,大部分对CCR5拮抗剂产生耐药的HIV?1突变株仍然保持原先的辅助受体嗜性。而且临床上有限的数据积累亦显示,已开发的CCR5拮抗剂在临床试验中还未发现有加快耐药株嗜性转换的病例。因此,CCR5拮抗剂的应用是不会加剧病情的恶化反应。目前,主要的CCR5受体拮抗剂有以下几种。 β趋化因子(RANTES,MIP?1α及MIP?1β)作为CCR5的天然配体是HIV?1受体当然的拮抗剂,在一定程度上可以保护细胞免受HIV?1的感染,其主要作用机制是诱导了CCR5的内吞作用(endocytosis)。然而,由于天然趋化因子具有半衰期短(<10min)以及潜在的炎症应答作用,它们作为拮抗剂药物并不是很合适。也有报道显示,高浓度的CC?趋化因子能够减缓病情的恶化,但是Marozsan等研究人员同时也发现高浓度的CC?趋化因子也可通过活化细胞而增强HIV?1的感染。而且Mosier等人发现CC?趋化因子也可促进R5嗜性病毒株向X4嗜性病毒株的转换以致病情加剧。因此与天然的配体相比,趋化因子衍生物通过不诱导信号通路而使受体内化的方式阻断相应的受体表位而更具优越性。
能治疗艾滋病的ccr5怎么查出来 。
怎么都是查艾滋病。提问者应该问的是怎么找到HIV和CCR5存在关系的。首先CCR5是趋化因子的一个蛋白质,是HIV结合蛋白。它在细胞表达,对其进行敲出导致HIV感染能力大大下降(不是没有,HIV有报道在有些情况下不需要结合CCR5诱导胞吞)来达到抑制艾滋病的目的。CCR5不是能治疗艾滋病的基因,趋化因子参与到天然免疫和先天免疫的过程中,有重要免疫作用,不能随便敲除。
CCR5怎么查出来的?
一般我们细胞表面的蛋白(MHC)成数量很多,不能可能一个个实验。为了找到病毒和哪一个蛋白存在作用,我们一般需要将病毒包膜蛋白(Env)表达纯化,并对Env制备成亲和柱,用亲和层析筛选MHC,最后对找到的MHC单独实验验证
CCR5的趋化因子
病毒特异的CD4+T细胞是针对HIV免疫应答的重要组成部分,也是HIV-1感染过程的首要靶点。自1984年发现HIV-1通过与CD4结合感染宿主细胞,随后研究又发现仅有CD4分子并不能介导HIV-1的侵入,同时还需要一种或几种辅助受体。1996年证实,趋化因子受体CXCR4和CCR5是HIV-1感染的辅助受体(coreceptor)[3,4]。而在2008年2月,美国国立卫生研究院又发现了另外一种新的HIV-1受体———整合素α4β7(integrinα4β7),此受体的发现也许可解释为什么消化道为HIV复制的主要场所,同时为艾滋病新药的开发指明方向[5]。
趋化因子是一类具有趋化活性的小分子细胞因子,它的功能在于使细胞表面具有跨膜G蛋白偶联受体的细胞聚集在一起,参与体内的免疫平衡作用。迄今为止,已发现人类趋化因子家族有50多个成员[6],根据其N?末端两个半胱氨酸残基的相对位置可分为4个不同结构的家族:CXC(或α)、CC(或β)、C(或γ)和CXC3C(或δ)。尽管CCS和CXCS具有相似的一级、二级和三级结构,但是由于两者四级结构的差异,它们分别作用于不同的受体[7]。趋化因子受体是属于鸟嘌呤核苷酸结合蛋白(G蛋白)偶联的7次跨膜蛋白受体家族,根据其配体可分为CXC和CC受体。迄今已鉴定出28种趋化因子受体,其中包括6种CXCRs(CXCR1~CXCR6)、10种CCRs(CCR1~CCR10)、1种XCR、1种CX3CR。其中的趋化因子受体CXCR4与CCR5,由于它们与HIV-1的侵染有关而备受关注。几乎所有的HIV-1病毒株都是利用其中的CCR5或CXCR4受体,或同时利用两种辅助受体侵入细胞的。有研究表明,CCR5和CXCR4的N-末端和第二胞外区对于它们与HIV-1gp120的结合起着关键的作用[8]。
利用何种趋化因子受体侵入细胞决定了HIV-1病毒株的嗜性。单核细胞/巨噬细胞嗜性的病毒株利用趋化因子受体CCR5侵入细胞,称为R5嗜性;T细胞嗜性的病毒株利用趋化因子受体CXCR4侵入细胞,称为X4嗜性;而既能利用CCR5,又能利用CXCR4侵入细胞的病毒株称为R5/X4嗜性。HIV-1gp120与何种辅助受体结合是随着HIV-1侵入靶细胞的不同阶段而有所变化的,通常在感染的初期是以CCR5为辅助受体;随着感染程度的加深,HIV-1由R5嗜性转化为R5/X4嗜性,而双嗜性的HIV-1是以CXCR4为主要辅助受体的。Cocchi等人研究预测,病毒包膜蛋白的V3环与HIV-1病毒株的嗜性转化有关,尤其是V3环中11位及25位的碱性氨基酸在不同类型病毒株中的差异可以说明一些问题。
布朗因为接受ccr5变异基因才治愈hiv科学家为何不用
布朗体内的一类特殊的免疫细胞已经被白血病清除干净,而其他艾滋病人则不具有相同的条件。
目前还没有医学技术可以完全实现这一定点“清除技术”。
原创文章,作者:聚禄鼎,如若转载,请注明出处:https://www.xxso.cn/151077.html